imported>Fiona E Graham |
|
| (50 intermediate revisions by 4 users not shown) |
| Line 1: |
Line 1: |
| {{CZ:(U00984) Appetite and Obesity, University of Edinburgh 2010/EZnotice}}
| |
| {{subpages}} | | {{subpages}} |
| | {{Image|Arc.JPG|right|600px|'''The SCN and orexigenic regions of the brain.''' SCN fibres terminate in and around the arcuate nucleus (ARC) and the ventral part of the lateral hypothalamus <ref name=Yi06>Yi CX (2006)Ventromedial arcuate nucleus communicates peripheral metabolic information to the suprachiasmatic nucleus. ''Endocrinology'' 147:283–94</ref>. Activation of the arcuate nucleus releases NYP and AgRP (two potent orexigenic peptides) into the PVN which ultimately stimulate feeding and slow metabolism <ref name=Schwartz00>Schwartz MW (2007)Central nervous system control of food intake ''Nature'' 404:661-671 </ref>. Adapted from Froy 2010 <ref name=Froy10/>}} |
|
| |
|
| The control of food intake is a flexible system whereby internal and external environmental cues can alter the timing of feeding and [[appetite]]. The [[suprachiasmatic nucleus]] in the [[hypothalamus]] is the vital coordinator of these stimuli that ultimately generates fluctuations in neuronal and hormonal activities that are known as ''[[circadian rhythms]]''. Circadian rhythms are driven by the daily variations in ambient light, which, by alterating gene expression, elicit a host of physiological responses including fluctuations in the hormones involved in appetite and food intake. Various factors such as temperature and social cues influence circadian rhythms; indeed, food intake itself can regulate these rhythms, but the neural mechanisms by which this occurs remains elusive. It has recently been proposed that there is a ‘food entrainable oscillator’ that exists independently of the SCN, and which controls food anticipation activity. The importance of circadian rhythms for ensuring good health has been highlighted by our modern day lifestyle, as [[jet-lag]] and shift work have shown that disruption of these delicate balances can lead to pathologies such as [[metabolic syndrome]].
| | The [[suprachiasmatic nucleus]] in the [[hypothalamus]] is the vital coordinator of the fluctuations in neuronal and hormonal activities that are known as ''[[circadian rhythms]]''. Circadian rhythms are driven by the daily variations in ambient light, which, by alterating gene expression, elicit a host of physiological responses including fluctuations in the hormones involved in [[appetite]]. Various factors such as temperature and social cues influence circadian rhythms; indeed, food intake itself can regulate these rhythms, but the neural mechanisms by which this occurs remains elusive. It has recently been proposed that there is a ‘food entrainable oscillator’ that is independent of the SCN, and which controls food anticipation activity. |
|
| |
|
| {{CZ:(U00984) Appetite and Obesity, University of Edinburgh 2010/EZnotice}}
| | ==The clock genes== |
| {{subpages}}
| | Clock mechanisms in biological cells arise from cycles of [[transcription]] and [[translation]] of [[clock genes]]. The clock genes regulate a rhythm in cells that, even without light, will maintain a roughly 24 hour rhythm. An extreme example is of cave fish which have evolved in complete darkness for millions of years and their clock genes are still present in their DNA.<ref name=Mendoza09>Mendoza ''et al.''(2009) Brain clocks: From the suprachiasmatic nuclei to a cerebral network. ''The Neuroscientist'' 15:5</ref> |
| | |
| The control of food intake is a flexible system whereby internal and external environmental cues can alter the timing of feeding and [[appetite]]. The [[suprachiasmatic nucleus]] in the [[hypothalamus]] is the vital coordinator of these stimuli that ultimately generates fluctuations in neuronal and hormonal activities that are known as ''[[circadian rhythms]]''. Circadian rhythms are driven by the daily variations in ambient light, which, by alterating gene expression, elicit a host of physiological responses including fluctuations in the hormones involved in appetite and food intake. Various factors such as temperature and social cues influence circadian rhythms; indeed, food intake itself can regulate these rhythms, but the neural mechanisms by which this occurs remains elusive. It has recently been proposed that there is a ‘food entrainable oscillator’ that exists independently of the SCN, and which controls food anticipation activity. The importance of circadian rhythms for ensuring good health has been highlighted by our modern day lifestyle, as [[jet-lag]] and shift work have shown that disruption of these delicate balances can lead to pathologies such as [[metabolic syndrome]].
| |
| | |
| ==The generation of Circadian Rhythms: the clock genes== | |
| | |
| Clock mechanisms are formed by the [[transcription]] and [[translation]] of [[clock genes]], which rely on feedback loops. In mammals, ''Clock'' and ''Bmal1'' genes are part of the positive loop and ''Per'' genes, the negative loop. The clock genes regulate a self-sustaining rhythm in cells that even without light will maintain a roughly 24 hour rhythm. An extreme example is of cave fish which have lived in complete darkness for millions of years and their clock genes are still present in their DNA.<ref name=Mendoza09>Mendoza ''et al.''(2009) Brain clocks: From the suprachiasmatic nuclei to a cerebral network. ''The Neuroscientist'' 15: 5 </ref> | |
| | |
| The rhythmic output of organs can be influenced by metabolic, endocrine and homeostatic events, as well as by the circadian clock. For example, the SCN can change the rhythm of expression of liver genes and enzymes without using clock genes, but through second messenger systems induced by the autonomic nervous system instead. Other genes can also affect circadian clock genes; for example the ''ROR-alpha'' gene is a positive regulator of ''Bmal1'', which regulates lipogenesis and lipid storage. <ref>(Lau ''et al.'' 2004)</ref>
| |
| | |
| Genes which encode important proteins of core clock mechanism are circadian locomotor output cycles kaput (''Clock''), brain and muscle-Arnt-like 1(''Bmal1''), ''Period1'' (''Per1''),''Period2'' (''Per2''), ''Period3'' (''Per3''),''Cryptochrome1''(''Cry1'') and ''Cryptochrome2''(''Cry2''). CLOCK (the protein product of ''Clock'') is a [[transcription factor]] which dimerises with BMAL1 (the protein product of ''Bmal1'') to activates transcription. CLOCK and BMAL1 are proteins which form basic helix-loop-helix transcription factor.Basic helix loop-helix is a structural motif.CLOCK and BMAL1 transcription factors form a heterodimeric complex. The above complex binds to E-box, which is a DNA sequence found in the promoter region of the gene and E-box like promoter sequences. The binding of CLOCK:BMAL1 complex to E-box located in the promoter region of ""Per"" and ""Cry"" genes and activates their transcription. PER and CRY proteins are able to undergo nuclear transformation and inhibit CLOCK:BMAL1 complex, which results in the decreased transcription of their owm genes. CLOCK:BMAL1 and PER-CRY transcription-translation loop is able to detect and self adjust to the changes. CLOCK:BMAL1 and PER-CRY constitute the core of the circadian clock and are able to generate 24 hour rhythms of gene expression.<ref>''Froy'' (2010)</ref>. Recent studies on the function and interaction of CLOCK:BMAL1 interaction have shown that it induces transcription in the following genes: RORɑ and REV-ERBɑ.<ref>''Froy'' (2010)</ref>. RORɑ and REV-ERBɑ regulate lipid metabolism and adipogenesis. The complex is also involved in the regulation of ""Bmal1"" transciption, with RORɑ stimulating and REV-ERBɑ inhibiting the transcription. Another gene which is activated by CLOCK:BMAL1 heterodimer is Pparɑ, which codes for a nuclear receptor involved in glucose and lipid metabolism.
| |
| | |
| Studies on mice with impaired ''Clock'' function showed increased food intake and eradication of rhythmic expression of ''Cart'' and ''Orexin'' hormones <ref>''Froy'' (2010)</ref>. Experimental data reported by Bray ''et.al'' shows that CLOCK -/- mice exhibit obesity,altered feeding patterns,hyperphagia and hormonal abnormalities similar to those found in metabolic syndromes, such as hyperlipidemia(abnormally elevated levels of lipoproteins in the blood), hyperglycemia()high amount of glucose in blood plasma) and hyperinsulinemia (excess levels of circulating insulin in the bloodstream).
| |
| | |
| ==The SCN, circadian rhythms and feeding behaviour==
| |
| | |
| A wide variety of organisms, from [[cyanobacteria]] to humans, all share common internal clock mechanisms that have been present for millions of years in evolutionary history. The circadian clock in mammals is responsible for setting specific temporal patterns within our bodily systems, including many physiological functions (body temperature, melatonin release, glucocorticoid secretion) and behavioural functions (alertness, working memory,) in order to keep us alive and running smoothly.
| |
| | |
| The "master" circadian clock of mammals is in the suprachiasmatic nucleus (SCN) of the hypothalamus. The main input to the SCN consists of signals encoding light; these are carried from the retina to the SCN by the retinohypothalamic tract. [[Vasoactive intestinal polypeptide]], released from a subpopulation of neurones in the SCN, activates and synchronises other SCN neurons, the output of which coordinates behavioural rhythms. One way in which the SCN may regulate food intake is by directing the sleep/wake cycle.
| |
| | |
| The role of the SCN in feeding patterns has been determined using rodents, as their food intake is less influenced by cognition and social behaviour as found in humans. The sleep/wake cycle is driven by fluctuations in two main hormones: [[corticosterone]] (cortisol in humans) and [[melatonin]]. In rats, corticosterone secretion increases during the night when these nocturnal animals are active. This rise is followed by an increase in locomotor activity, in which rats forage for food and subsequently begin to feed. Melatonin is an important hormone that is released from the [[pineal gland]] during the night which, amongst its many actions affects sleep and appetite.
| |
| | |
| | |
| | |
| | |
| {{Image|SCN.JPG|centre|700px|.'''Figure 2. SCN regulation of the sleep/wake cycle''';
| |
| The SCN innervates multiple regions of the brain that regulate the sleep-wake cycle. a) The most intense output from the SCN is to the ventral sub-paraventricular zone (SPZ); this projects to the dorsomedial hypothalamus (DMH) which controls a wide range of circadian responses including corticosteroid secretion. The DMH innervates the medial parvicellular paraventricular nucleus (PVHm) and regulates the CRH neurons which control pituitary regulation of corticosteroid production. The SCN is also involved in regulating the release of melatonin from the pineal gland. The SCN projects to the dorsal parvicellular paraventricular nucleus, (PVHd) which projects to sympathetic preganglionic neurons in the spinal cord which regulate melatonin output by the release of noradrenaline into the pineal gland. b) The DMH also has GABAergic projections to the ventrolateral preoptic nucleus VLPO (the sleep promoting region) and glutamatergic projections to the orexin producing neurons and the melanin-concentrating hormone (MCH)neurons in the lateral hypothalamus (LHA) which together regulate sleep and wakefulness and ultimately feeding. Hormones involved in appetite regulation such as ghrelin and leptin can influence these areas of the brain thus may have a role in resetting the circadian rhythms generated by the SCN. (Adapted from figure 2c&d Saper ''et al.'' (2005)<ref>Saper A ''et al.'' (2005) The hypothalamic intergrator for circadian rhythms ''Trends Neurosci'' 28:152-157</ref>.) }}
| |
| | |
| | |
| However, it is important to note that these hormones have different physiological roles in different species. For example, melatonin is released during the night in both humans and rodents, and yet latter are nocturnal, thus are most active at night. This indicates that melatonin has the opposite effects in rodents as it does in humans in that it appears to drive the awake period in rats and induce the sleep period in humans. Thus it is appearent that whilst the SCN may regulate the sleep-wake cycle, this regulation is species-specific which suggests that the role the SCN has in feeding behaviours is equally complex due to the diverse actions of the hormones involved.
| |
| | |
| Nevertheless, it is possible that that the SCN may drive feeding patterns directly as the SCN appears to have reciprocal interactions with the orexigenic regions of the brain namely the lateral hypothalamus.
| |
| | |
| {{Image|Arc.JPG|centre|500px|'''Figure 3. The SCN and orexigneic regions of the brain.''' SCN fibres terminate in and around the arcuate nucleus (ARC) and the ventral part of the lateral hypothalamus <ref>Yi ''et al.'' (2006)</ref>. Activation of the arcuate nucleus releases NYP and AgRP (two potent orexigenic peptides) into the PVN which ultimately stimulates feeding and slows metabolism <ref name=Schwartz00>Schwartz MW (2007)Central nervous system control of food intake ''Nature'' 404:661-671 </ref>. Adapted from Fig 3. Froy 2009}}
| |
| | |
| | |
| Alternatively, it is possible that the SCN initiates feeding by conducting circadian rhythmic oscillations in the hormones involved in appetite. Indeed, a number of hormones involved in feeding behaviour and appetite, including leptin and ghrelin, show circadian oscillations. <ref name=Yildiz04>Yildiz BO (2004) Alteration in the dynamics of circulating ghrelin, adiponectin and leptin in human obesity ''Proc Natl Acad Sci USA'' 101:10434-9</ref>. Leptin has been shown to exhibit circadian patterns in both gene expression and protein secretion in humans, with a peak during the sleep phase in humans (Kalra ''et al.'' 2003 in froy). Furthermore, rodent studies have shown that ablation of the SCN eliminates leptin circadian rhythmicity (Kalsbeek 2001 in Froy) and yet the role of the SCN in conducting this pattern is unclear. As leptin binds to receptors in the hypothalamus to suppress of appetite and an increase metabolism <ref name=Schwartz00/> it seems plausible to suggest that the SCN can alter appetite indirectly via hormone regulation.
| |
| | |
| Whilst the role the SCN has in driving feeding behaviour remains elusive the latest research indicates that in addition to this central circadian clock there exists peripheral oscillators in other areas of the brain and other organs that produce rhythmic patterns in feeding time. Furthermore, it has been shown that rodents show food anticipatory behaviour indicating that these peripheral oscillators can be reset by feeding time itself thus have been named food entrainable oscillators. Moreover, it has been suggested that the SCN can signal to peripheral oscillators using signalling molecules such as TGF-alpha and prokinecticin 2 that prevent dampening of circadian rhythms in the tissues FROY. The recent study by Gilbert ''et al.'' (2009) have shown that transforming growth factor alpha is an important SCN peptide that regulates activity and feeding behaviour. Indeed intraperitoneal infusions of TGF-alpha was found to not only reduce locomotor activity but also to cause reduced food consumption and weight loss in Syrian hamsters <ref name=Gilbert09>Gilbert J (2009) Behavioral effects of systemic transforming growth factor-alpha in Syrian hamsters. ''Behav Brain Res'' 198:440-448</ref> Whilst the site of action of TGF alpha in this experiment was found to be the brain it is also possible that they act outside the central nervous system.
| |
| | |
| [[User:Fiona E Graham|Fiona E Graham]] 15:57, 25 October 2010 (UTC)
| |
| | |
| ==Peripheral Clocks and Food Entrainable Oscillators==
| |
| | |
| :'''Peripheral clocks'''
| |
| | |
| | |
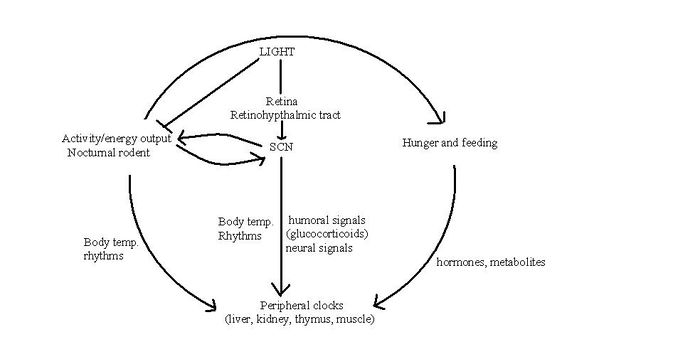
| The SCN and peripheral clocks are not affected by meal timings, but restriction of food is the dominant synchroniser for peripheral clocks. When rats are restricted of food, metabolic and hormonal factors used by the SCN to drive peripheral oscillators are uncoupled, to shift to food time. Normally, the SCN and peripheral oscillators will work together as one unit, but a change in food availability can uncouple them in order for survival, when feeding is low and shifted from their normal place in the light-dark cycle.<ref>Escobar C ''et al.'' (2009) Peripheral oscillators are important for food anticipatory activity (FAA)European Journal of Neuroscience, 30: 1665–1675 '''' </ref>.
| |
| | |
| {{Image|Circ rhythm.JPG|right|350px|SCN and Peripheral Oscillator Interaction.}}
| |
| | |
| :'''The Food Entrained Oscillator'''
| |
| | |
| The FEO is a mysterious circadian clock, which is independent of the SCN. It ensures that when food is scarce, the body is still ready to digest and extract nutrients from the food that has been found, the FEO is responsible for anticipation of meal-time (FAA) <ref name=Mendoza09/>.
| |
| | |
| Clock genes may contribute to the FAA but are not essential – Pendergast et al (2009) showed that animals without essential clock genes Bmal1 or per1 or2 were arrhythmic in constant darkness but still could express FAA.
| |
| The FEO’s existence is putative, and its network is believed to be scattered over several brain regions. For example, the dorsomedial hypothalamic nucleus has been reported for FAA expression and the possible site of the FEO. The circadian mechanism for the FEO is unknown, but it does present clear circadian features.
| |
| | |
| [[User:Laura Sheldon|Laura Sheldon]] 19:09, 24 October 2010 (UTC)Laura
| |
| | |
| ==Sleep deprivation, shift-work and appetite==
| |
|
| |
|
| {{Image|Fig 1.PNG|right|350px|''' The relationship between sleep duration and changes in serum leptin and ghrelin levels.''' '''(a)''' Mean leptin levels against average nightly sleep duration. As the number of hours sleep increases, the levels of serum leptin also increase. Standard errors for half-hour increments of average nightly sleep. '''(b)''' Mean ghrelin levels against total number of hours sleep. As the total number of hours sleep decreases, the mean levels of ghrelin increase. Standard errors for half-hour increments of total sleep time. Adapted from Taheri ''et. al'' (2004)}}
| | Genes which encode important proteins of the core clock mechanism include ''Clock'' (circadian locomotor output cycles kaput); ''Bmal1'' (brain and muscle-Arnt-like 1); the ''Period'' genes ''Per1'', ''Per2'' and ''Per3''); and the ''Cryptochrome'' genes ''Cry1'' and ''Cry2''. CLOCK (the protein product of ''Clock'') is a [[transcription factor]] which dimerises with BMAL1 (the protein product of ''Bmal1''). CLOCK and BMAL1 form a complex which binds to E-box, a DNA sequence in the promoter region of the gene, and to other similar promoter sequences. The binding of the CLOCK:BMAL1 complex to the E-box in the promoter region of ""Per"" and ""Cry"" activates their transcription. In turn, the PER and CRY proteins can inhibit the CLOCK:BMAL1 complex, resulting in the decreased transcription of their own genes.<ref name=Froy10>Froy ''et al.''(2010) Metabolism and circadian rhythms—implications for obesity ''Endocr Rev'' 31:1-24</ref> |
|
| |
|
| In modern society, where shopping, eating, working and drinking are widely available 24hours a day,major health implications have been linked as a result. This availability of around the clock activities has defied our bodies internal clock of the vital hours of sleep it requires <ref>Gimble JM ''et al.'' (2009) Circadian biology and sleep: missing links in obesity and metabolism? ''Obesity Rev'' 10(suppl 2):1-5.</ref>. Over the last few decades where technology and social activities has dramatically advanced, the number of hours of sleep young adults get has decreased within the range of 1-2hours <ref name=Spiegel04>Spiegel ''et. al''(2004)Brief communication: Sleep curtailment in healthy young men is associated with decreased leptin levels, elevated ghrelin levels, and increased hunger and appetite ''Ann Internal Med'' 141:846-50</ref>. This is strongly correlated with the prevalence of obesity within the U.K which has shown to have trebled over the last 3 decades <ref>Rennie KL, Jebb SA (2005) Prevalence of obesity in Great Britain ''Obesity Rev'' 6:11-2</ref>. Although the rise in this obesity epidemic has been shown to be multi-factorial, sleep deprivation is just another factor to add to the list needing to be addressed in this ever rising health problem.
| | Mice with impaired ''Clock'' function are obese, displaying altered feeding patterns, hyperphagia and abnormalities similar to those found in metabolic syndromes, such as hyperlipidemia (abnormally elevated levels of lipoproteins in the blood), hyperglycemia (high concentrations of glucose in blood) and hyperinsulinemia (excess of circulating insulin).<ref name=Froy10/> |
|
| |
| Several epidemiologic studies have shown that sleep deprivation elevates the levels of the appetite stimulating hormone [[ghrelin]] and decreases circulating [[leptin]] levels [Fig. 5.1] <ref name=Taheri04> Taheri ''et al.'' (2004) Short sleep duration is associated with reduced leptin, elevated ghrelin, and increased body mass index. ''PLoS Medicine'' 1(3):e62</ref><ref name=Spiegel04/>. These changes could be the causes of increased food intake in these sleep- deprived adults where a rise in body weight is also observed [Fig. 5.2] <ref name=Taheri04/>. It is therefore important to fully understand the health implications that sleep-deprivation, associated with jetlag and night shift workers, has on the body in order to develop future therapeutic schemes against such related disorders as obesity.
| |
| | |
| {{Image|Fig2.PNG|centre|350px|'''The relationship between average nightly sleep and changes in body mass index (BMI).''' As the average nightly hours of sleep decreases below 7 hours, the mean BMI increases. Furthermore, more than an average of 8 hours sleep also causes a rise in mean BMI. Standard errors for 45-min intervals of average nightly sleep. Adapted from Taheri ''et. al'' (2004).}}
| |
| | |
| ==Conclusion==
| |
| | |
| Over the last decade, significant progress has been made in our understanding of the inter-relationship between circadian biology, sleep and metabolism in the context of obesity. The SCN which is a vital coordinator of circadian rhythms may have a possible role for driving feeding patterns, along with other peripheral oscillators in the brain and organs that produce rhythmic patterns in feeding time.
| |
| | |
| As a result of this newly industrialized world, sleep deprivation is a growing issue of concern and the serious health implications it has on the population need to be acknowledged. An adequate number of hours sleep each night is vital in ensuring a healthy lifestyle. Disruptions of the circadian variations in such appetite-regulating hormones as leptin and ghrelin may contribute to the development of obesity in sleep deprived patients.
| |
| | |
| Gaps still remain in our knowledge of the underlying biological rhythms and its association with the pathogenesis of obesity. So far previous therapeutic strategies have failed to curb the rising incidence of obesity, but if, through a greater understanding of these complex systems we are able to manipulate and control appetite the benefits will be a healthier population and less strain on the health services caused by obesity.
| |
| | |
| ==References==
| |
| <references/>
| |
|
| |
|
| ==The SCN, circadian rhythms and feeding behaviour== | | ==The SCN, circadian rhythms and feeding behaviour== |
| | {{Image|SCN.JPG|left|600px|'''SCN regulation of the sleep/wake cycle'''; |
| | The SCN innervates regions of the brain that regulate the sleep-wake cycle. a) The ventral sub-paraventricular zone (SPZ) projects to the [[dorsomedial hypothalamus]] (DMH) which controls many circadian responses including [[corticosteroid]] secretion. The DMH innervates the medial parvicellular [[paraventricular nucleus]] (PVHm) and regulates the neurons which control corticosteroid production. The SCN also projects to the dorsal parvicellular paraventricular nucleus, (PVHd) which projects to sympathetic preganglionic neurons which regulate [[melatonin]] output from the [[pineal gland]]. b) The DMH also has projections to the ventrolateral preoptic nucleus VLPO (the sleep promoting region) and to the [[orexin]] neurons and [[melanin-concentrating hormone]] (MCH) neurons in the [[lateral hypothalamus]] (LHA) which together regulate sleep and wakefulness. Hormones involved in appetite regulation such as ghrelin and leptin can influence these areas of the brain thus may have a role in resetting the circadian rhythms generated by the SCN. (Adapted from Saper ''et al.'' (2005)<ref>Saper A ''et al.'' (2005) The hypothalamic integrator for circadian rhythms ''Trends Neurosci'' 28:152-7</ref>)}} |
|
| |
|
| A wide variety of organisms, from [[cyanobacteria]] to humans, all share common internal clock mechanisms that have been present for millions of years in evolutionary history. The circadian clock in mammals is responsible for setting specific temporal patterns within our bodily systems, including many physiological functions (body temperature, melatonin release, glucocorticoid secretion) and behavioural functions (alertness, working memory,) in order to keep us alive and running smoothly.
| | The circadian clock in mammals sets specific temporal patterns within our bodily systems, including many physiological functions (body temperature, melatonin release, glucocorticoid secretion) and behavioural functions. The "master" circadian clock of mammals is in the [[suprachiasmatic nucleus]] (SCN) of the hypothalamus. This receives signals encoding light, which are carried from the retina to the SCN by the retinohypothalamic tract. [[Vasoactive intestinal polypeptide]], [[vasopressin]], [[somatostatin]] and other peptides, released from subpopulations of neurones in the SCN, activate and synchronise other SCN neurons, the output of which coordinates behavioural rhythms, including by directing the sleep/wake cycle. The sleep/wake cycle is driven by fluctuations in two main hormones: [[corticosterone]] (cortisol in humans) and [[melatonin]]. In rats, [[corticosterone]] secretion increases during the night when they are active. This rise is followed by an increase in locomotor activity, in which rats forage for food and subsequently begin to feed. [[Melatonin]] is an important hormone that is released from the [[pineal gland]] during the night and, amongst its many actions, it affects sleep and appetite. However, these hormones have different roles in different species. For example, melatonin is released during the night in both humans and rodents, and yet latter are nocturnal and therefore most active at night. This indicates that melatonin has opposite effects in rodents and humans in that it appears to drive the awake period in rats, but induces sleep in humans. Thus whilst the SCN may regulate the sleep-wake cycle, this regulation is species-specific. Nevertheless, it is possible that the SCN may drive feeding patterns directly as the SCN has reciprocal interactions with orexigenic regions of the brain such as the lateral hypothalamus. |
| | |
| The "master" circadian clock of mammals is in the suprachiasmatic nucleus (SCN) of the hypothalamus. The main input to the SCN consists of signals encoding light; these are carried from the retina to the SCN by the retinohypothalamic tract. [[Vasoactive intestinal polypeptide]], released from a subpopulation of neurones in the SCN, activates and synchronises other SCN neurons, the output of which coordinates behavioural rhythms. One way in which the SCN may regulate food intake is by directing the sleep/wake cycle. | |
| | |
| The role of the SCN in feeding patterns has been determined using rodents, as their food intake is less influenced by cognition and social behaviour as found in humans. The sleep/wake cycle is driven by fluctuations in two main hormones: [[corticosterone]] (cortisol in humans) and [[melatonin]]. In rats, corticosterone secretion increases during the night when these nocturnal animals are active. This rise is followed by an increase in locomotor activity, in which rats forage for food and subsequently begin to feed. Melatonin is an important hormone that is released from the [[pineal gland]] during the night which, amongst its many actions affects sleep and appetite.
| |
| | |
| | |
| | |
| | |
| {{Image|SCN.JPG|centre|700px|.'''Figure 2. SCN regulation of the sleep/wake cycle''';
| |
| The SCN innervates multiple regions of the brain that regulate the sleep-wake cycle. a) The most intense output from the SCN is to the ventral sub-paraventricular zone (SPZ); this projects to the dorsomedial hypothalamus (DMH) which controls a wide range of circadian responses including corticosteroid secretion. The DMH innervates the medial parvicellular paraventricular nucleus (PVHm) and regulates the CRH neurons which control pituitary regulation of corticosteroid production. The SCN is also involved in regulating the release of melatonin from the pineal gland. The SCN projects to the dorsal parvicellular paraventricular nucleus, (PVHd) which projects to sympathetic preganglionic neurons in the spinal cord which regulate melatonin output by the release of noradrenaline into the pineal gland. b) The DMH also has GABAergic projections to the ventrolateral preoptic nucleus VLPO (the sleep promoting region) and glutamatergic projections to the orexin producing neurons and the melanin-concentrating hormone (MCH)neurons in the lateral hypothalamus (LHA) which together regulate sleep and wakefulness and ultimately feeding. Hormones involved in appetite regulation such as ghrelin and leptin can influence these areas of the brain thus may have a role in resetting the circadian rhythms generated by the SCN. (Adapted from figure 2c&d Saper ''et al.'' (2005)<ref>Saper A ''et al.'' (2005) The hypothalamic intergrator for circadian rhythms ''Trends Neurosci'' 28:152-157</ref>.) }}
| |
| | |
| | |
| However, it is important to note that these hormones have different physiological roles in different species. For example, melatonin is released during the night in both humans and rodents, and yet latter are nocturnal, thus are most active at night. This indicates that melatonin has the opposite effects in rodents as it does in humans in that it appears to drive the awake period in rats and induce the sleep period in humans. Thus it is appearent that whilst the SCN may regulate the sleep-wake cycle, this regulation is species-specific which suggests that the role the SCN has in feeding behaviours is equally complex due to the diverse actions of the hormones involved. | |
|
| |
|
| Nevertheless, it is possible that that the SCN may drive feeding patterns directly as the SCN appears to have reciprocal interactions with the orexigenic regions of the brain namely the lateral hypothalamus.
| | Alternatively, the SCN may initiate feeding by conducting circadian rhythmic oscillations in the hormones involved in appetite. Indeed, a number of hormones involved in feeding behaviour and appetite, including leptin and ghrelin, show circadian oscillations. <ref name=Yildiz04>Yildiz BO (2004) Alteration in the dynamics of circulating [[ghrelin]], [[adiponectin]] and [[leptin]] in human obesity ''Proc Natl Acad Sci USA'' 101:10434-9</ref>. Leptin exhibits circadian patterns in both gene expression and protein secretion in humans, with a peak during sleep <ref name=Kalra03>Kalra SP(2003) Rhythmic, reciprocal ghrelin and leptin signaling: new insight in the development of obesity ''Regul Pept'' 111:1–11</ref>. In rodents, ablation of the SCN eliminates leptin circadian rhythmicity <ref name=Kalsbeek01>Kalsbeek A (2001) The suprachiasmatic nucleus generates the diurnal changes in plasma leptin levels ''Endocrinology'' 142:2677–85</ref> and yet the role of the SCN in conducting this pattern is unclear. |
|
| |
|
| {{Image|Arc.JPG|centre|500px|Adapted from Fig 3. Froy 2009. SCN fibres terminate in and around the arcuate nucleus (ARC) and the ventral part of the lateral hypothalamus <ref>Yi ''et al.'' (2006)</ref>. Activation of the arcuate nucleus releases NYP and AgRP (two potent orexigenic peptides) into the PVN which ultimately stimulates feeding and slows metabolism <ref name=Schwartz00>Schwartz MW (2007)Central nervous system control of food intake ''Nature'' 404:661-671 </ref>.}}
| | Whilst the role the SCN has in driving feeding behaviour remains elusive, oscillators in other areas of the brain and other organs may also produce rhythmic patterns in feeding. Rodents show ''food anticipatory behaviour'' indicating that these peripheral oscillators can be reset by feeding time itself, thus have been named 'food entrainable oscillators'. It has been suggested that the SCN signals to these other oscillators using signalling molecules such as [[transforming growth factor alpha]] (TGF-alpha) and prokinecticin 2 that prevent dampening of circadian rhythms in the tissues <ref name=Froy10/>. <ref name=Gilbert09>Gilbert J (2009) Behavioral effects of systemic transforming growth factor-alpha in Syrian hamsters ''Behav Brain Res'' 198:440-8</ref> |
| | |
| | |
| Alternatively, it is possible that the SCN initiates feeding by conducting circadian rhythmic oscillations in the hormones involved in appetite. Indeed, a number of hormones involved in feeding behaviour and appetite, including leptin and ghrelin, show circadian oscillations. <ref name=Yildiz04>Yildiz BO (2004) Alteration in the dynamics of circulating ghrelin, adiponectin and leptin in human obesity ''Proc Natl Acad Sci USA'' 101:10434-9</ref>. Leptin has been shown to exhibit circadian patterns in both gene expression and protein secretion in humans, with a peak during the sleep phase in humans (Kalra ''et al.'' 2003 in froy). Furthermore, rodent studies have shown that ablation of the SCN eliminates leptin circadian rhythmicity (Kalsbeek 2001 in Froy) and yet the role of the SCN in conducting this pattern is unclear. As leptin binds to receptors in the hypothalamus to suppress of appetite and an increase metabolism <ref name=Schwartz00/> it seems plausible to suggest that the SCN can alter appetite indirectly via hormone regulation.
| |
| | |
| Whilst the role the SCN has in driving feeding behaviour remains elusive the latest research indicates that in addition to this central circadian clock there exists peripheral oscillators in other areas of the brain and other organs that produce rhythmic patterns in feeding time. Furthermore, it has been shown that rodents show food anticipatory behaviour indicating that these peripheral oscillators can be reset by feeding time itself thus have been named food entrainable oscillators. Moreover, it has been suggested that the SCN can signal to peripheral oscillators using signalling molecules such as TGF-alpha and prokinecticin 2 that prevent dampening of circadian rhythms in the tissues FROY. The recent study by Gilbert ''et al.'' (2009) have shown that transforming growth factor alpha is an important SCN peptide that regulates activity and feeding behaviour. Indeed intraperitoneal infusions of TGF-alpha was found to not only reduce locomotor activity but also to cause reduced food consumption and weight loss in Syrian hamsters <ref name=Gilbert09>Gilbert J (2009) Behavioral effects of systemic transforming growth factor-alpha in Syrian hamsters. ''Behav Brain Res'' 198:440-448</ref> Whilst the site of action of TGF alpha in this experiment was found to be the brain it is also possible that they act outside the central nervous system. | |
| | |
| [[User:Fiona E Graham|Fiona E Graham]] 15:57, 25 October 2010 (UTC)
| |
|
| |
|
| ==Peripheral Clocks and Food Entrainable Oscillators== | | ==Peripheral Clocks and Food Entrainable Oscillators== |
| | {{Image|Circ rhythm.JPG|right|700px|SCN and Peripheral Oscillator Interaction.}} |
| | The SCN and peripheral clocks are not affected by meal timings, but restriction of food is the dominant synchroniser for peripheral clocks. When rats are restricted of food, metabolic and hormonal factors used by the SCN to drive peripheral oscillators are uncoupled, to shift to food time. Normally, the SCN and peripheral oscillators work together, but a change in food availability can uncouple them when environmental circumstances demand that feeding patterns are shifted from their normal place in the light-dark cycle.<ref>Escobar C ''et al.'' (2009) Peripheral oscillators are important for food anticipatory activity (FAA) ''Eur J Neurosci'' 30:1665–75</ref>. |
|
| |
|
| :'''Peripheral clocks'''
| | The 'Food Entrained Oscillator' is a mysterious circadian clock, which is independent of the SCN. It ensures the body is ready to digest and extract nutrients from the food in anticipation of meal-time <ref name=Mendoza09/>. Clock genes may contribute to the Food Entrained Oscillator, but are not essential – animals without essential clock genes ''Bmal1'' or ''per1'' or ''per2'' were arrhythmic in constant darkness but still could express food anticipation. <ref>Pendergast ''et al''' (2009)</ref> |
| | |
| | |
| The SCN and peripheral clocks are not affected by meal timings, but restriction of food is the dominant synchroniser for peripheral clocks. When rats are restricted of food, metabolic and hormonal factors used by the SCN to drive peripheral oscillators are uncoupled, to shift to food time. Normally, the SCN and peripheral oscillators will work together as one unit, but a change in food availability can uncouple them in order for survival, when feeding is low and shifted from their normal place in the light-dark cycle.<ref>Escobar C ''et al.'' (2009) Peripheral oscillators are important for food anticipatory activity (FAA)European Journal of Neuroscience, 30: 1665–1675 '''' </ref>. | |
| | |
| {{Image|Circ rhythm.JPG|right|350px|SCN and Peripheral Oscillator Interaction.}}
| |
| | |
| :'''The Food Entrained Oscillator'''
| |
| | |
| The FEO is a mysterious circadian clock, which is independent of the SCN. It ensures that when food is scarce, the body is still ready to digest and extract nutrients from the food that has been found, the FEO is responsible for anticipation of meal-time (FAA) <ref name=Mendoza09/>.
| |
| | |
| Clock genes may contribute to the FAA but are not essential – Pendergast et al (2009) showed that animals without essential clock genes Bmal1 or per1 or2 were arrhythmic in constant darkness but still could express FAA. | |
| The FEO’s existence is putative, and its network is believed to be scattered over several brain regions. For example, the dorsomedial hypothalamic nucleus has been reported for FAA expression and the possible site of the FEO. The circadian mechanism for the FEO is unknown, but it does present clear circadian features.
| |
| | |
| [[User:Laura Sheldon|Laura Sheldon]] 19:09, 24 October 2010 (UTC)Laura
| |
|
| |
|
| ==Sleep deprivation, shift-work and appetite== | | ==Sleep deprivation, shift-work and appetite== |
| | In modern society, shopping, eating, working and drinking are widely available 24 hours a day. Over the last few decades, the number of hours of sleep young adults get has decreased by 1-2 hours. This is correlated with the prevalence of obesity within the U.K which has trebled over the last three decades. Although the rise in this obesity epidemic is multi-factorial, sleep deprivation may be another factor to add to the list needing to be addressed in this ever rising health problem. <ref>Gimble JM ''et al.'' (2009) Circadian biology and sleep: missing links in obesity and metabolism? ''Obesity Rev'' 10(suppl 2):1-5</ref> <ref>Rennie KL, Jebb SA (2005) Prevalence of obesity in Great Britain ''Obesity Rev'' 6:11-2</ref> <ref name=Spiegel04>Spiegel ''et al.''(2004) Sleep curtailment in healthy young men is associated with decreased leptin levels, elevated ghrelin levels, and increased hunger and appetite ''Ann Internal Med'' 141:846-50</ref>. |
|
| |
|
| {{Image|Fig 1.PNG|right|350px|''' The relationship between sleep duration and changes in serum leptin and ghrelin levels.''' '''(a)''' Mean leptin levels against average nightly sleep duration. As the number of hours sleep increases, the levels of serum leptin also increase. Standard errors for half-hour increments of average nightly sleep. '''(b)''' Mean ghrelin levels against total number of hours sleep. As the total number of hours sleep decreases, the mean levels of ghrelin increase. Standard errors for half-hour increments of total sleep time. Adapted from Taheri ''et. al'' (2004)}}
| | Sleep deprivation elevates the circulating levels of the appetite stimulating hormone [[ghrelin]] and decreases those of [[leptin]]<ref name=Taheri04> Taheri ''et al.'' (2004) Short sleep duration is associated with reduced leptin, elevated ghrelin, and increased body mass index ''PLoS Medicine'' 1:e62</ref><ref name=Spiegel04/>. These changes could be the causes of increased food intake in those sleep-deprived adults where a rise in body weight is also observed <ref name=Taheri04/>. It is therefore important to understand the health implications that sleep-deprivation, associated with jetlag and shift working, has on the body in order to develop therapeutic schemes against such related disorders as obesity. |
| | |
| In modern society, where shopping, eating, working and drinking are widely available 24hours a day,major health implications have been linked as a result. This availability of around the clock activities has defied our bodies internal clock of the vital hours of sleep it requires <ref>Gimble JM ''et al.'' (2009) Circadian biology and sleep: missing links in obesity and metabolism? ''Obesity Rev'' 10(suppl 2):1-5.</ref>. Over the last few decades where technology and social activities has dramatically advanced, the number of hours of sleep young adults get has decreased within the range of 1-2hours <ref name=Spiegel04>Spiegel ''et. al''(2004)Brief communication: Sleep curtailment in healthy young men is associated with decreased leptin levels, elevated ghrelin levels, and increased hunger and appetite ''Ann Internal Med'' 141:846-50</ref>. This is strongly correlated with the prevalence of obesity within the U.K which has shown to have trebled over the last 3 decades <ref>Rennie KL, Jebb SA (2005) Prevalence of obesity in Great Britain ''Obesity Rev'' 6:11-2</ref>. Although the rise in this obesity epidemic has been shown to be multi-factorial, sleep deprivation is just another factor to add to the list needing to be addressed in this ever rising health problem.
| |
|
| |
| Several epidemiologic studies have shown that sleep deprivation elevates the levels of the appetite stimulating hormone [[ghrelin]] and decreases circulating [[leptin]] levels [Fig. 5.1] <ref name=Taheri04> Taheri ''et al.'' (2004) Short sleep duration is associated with reduced leptin, elevated ghrelin, and increased body mass index. ''PLoS Medicine'' 1(3):e62</ref><ref name=Spiegel04/>. These changes could be the causes of increased food intake in these sleep- deprived adults where a rise in body weight is also observed [Fig. 5.2] <ref name=Taheri04/>. It is therefore important to fully understand the health implications that sleep-deprivation, associated with jetlag and night shift workers, has on the body in order to develop future therapeutic schemes against such related disorders as obesity.
| |
| | |
| {{Image|Fig2.PNG|centre|350px|'''The relationship between average nightly sleep and changes in body mass index (BMI).''' As the average nightly hours of sleep decreases below 7 hours, the mean BMI increases. Furthermore, more than an average of 8 hours sleep also causes a rise in mean BMI. Standard errors for 45-min intervals of average nightly sleep. Adapted from Taheri ''et. al'' (2004).}}
| |
|
| |
|
| ==Conclusion== | | ==Conclusion== |
| | | Over the last decade, significant progress has been made in our understanding of the inter-relationship between circadian biology, sleep and metabolism in the context of obesity. The SCN, which is a vital coordinator of circadian rhythms, may have a role in driving feeding patterns, along with other peripheral oscillators in the brain and organs that produce rhythmic patterns in feeding time. In the modern world, sleep deprivation is a growing issue of concern and the health implications need to be acknowledged. An adequate number of hours sleep each night is vital for a healthy life. Disruptions of the circadian variations in such appetite-regulating hormones as leptin and ghrelin may contribute to the development of obesity in sleep-deprived patients. Gaps still remain in our knowledge of the underlying biological rhythms and its association with obesity. So far, therapeutic strategies have failed to curb the rising incidence of obesity, but if, through a greater understanding of these complex systems we can control appetite, the benefits will be a healthier population and less strain on the health services. |
| Over the last decade, significant progress has been made in our understanding of the inter-relationship between circadian biology, sleep and metabolism in the context of obesity. The SCN which is a vital coordinator of circadian rhythms may have a possible role for driving feeding patterns, along with other peripheral oscillators in the brain and organs that produce rhythmic patterns in feeding time. | |
| | |
| As a result of this newly industrialized world, sleep deprivation is a growing issue of concern and the serious health implications it has on the population need to be acknowledged. An adequate number of hours sleep each night is vital in ensuring a healthy lifestyle. Disruptions of the circadian variations in such appetite-regulating hormones as leptin and ghrelin may contribute to the development of obesity in sleep deprived patients.
| |
| | |
| Gaps still remain in our knowledge of the underlying biological rhythms and its association with the pathogenesis of obesity. So far previous therapeutic strategies have failed to curb the rising incidence of obesity, but if, through a greater understanding of these complex systems we are able to manipulate and control appetite the benefits will be a healthier population and less strain on the health services caused by obesity. | |
|
| |
|
| ==References== | | ==References== |
| <references/>
| | {{reflist|2}}[[Category:Suggestion Bot Tag]] |
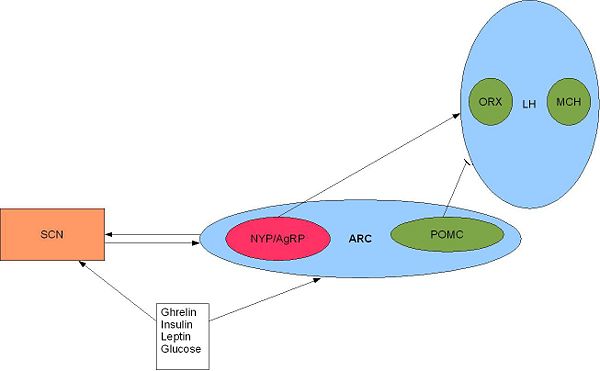
The SCN and orexigenic regions of the brain. SCN fibres terminate in and around the arcuate nucleus (ARC) and the ventral part of the lateral hypothalamus
[1]. Activation of the arcuate nucleus releases NYP and AgRP (two potent orexigenic peptides) into the PVN which ultimately stimulate feeding and slow metabolism
[2]. Adapted from Froy 2010
[3] The suprachiasmatic nucleus in the hypothalamus is the vital coordinator of the fluctuations in neuronal and hormonal activities that are known as circadian rhythms. Circadian rhythms are driven by the daily variations in ambient light, which, by alterating gene expression, elicit a host of physiological responses including fluctuations in the hormones involved in appetite. Various factors such as temperature and social cues influence circadian rhythms; indeed, food intake itself can regulate these rhythms, but the neural mechanisms by which this occurs remains elusive. It has recently been proposed that there is a ‘food entrainable oscillator’ that is independent of the SCN, and which controls food anticipation activity.
The clock genes
Clock mechanisms in biological cells arise from cycles of transcription and translation of clock genes. The clock genes regulate a rhythm in cells that, even without light, will maintain a roughly 24 hour rhythm. An extreme example is of cave fish which have evolved in complete darkness for millions of years and their clock genes are still present in their DNA.[4]
Genes which encode important proteins of the core clock mechanism include Clock (circadian locomotor output cycles kaput); Bmal1 (brain and muscle-Arnt-like 1); the Period genes Per1, Per2 and Per3); and the Cryptochrome genes Cry1 and Cry2. CLOCK (the protein product of Clock) is a transcription factor which dimerises with BMAL1 (the protein product of Bmal1). CLOCK and BMAL1 form a complex which binds to E-box, a DNA sequence in the promoter region of the gene, and to other similar promoter sequences. The binding of the CLOCK:BMAL1 complex to the E-box in the promoter region of ""Per"" and ""Cry"" activates their transcription. In turn, the PER and CRY proteins can inhibit the CLOCK:BMAL1 complex, resulting in the decreased transcription of their own genes.[3]
Mice with impaired Clock function are obese, displaying altered feeding patterns, hyperphagia and abnormalities similar to those found in metabolic syndromes, such as hyperlipidemia (abnormally elevated levels of lipoproteins in the blood), hyperglycemia (high concentrations of glucose in blood) and hyperinsulinemia (excess of circulating insulin).[3]
The SCN, circadian rhythms and feeding behaviour
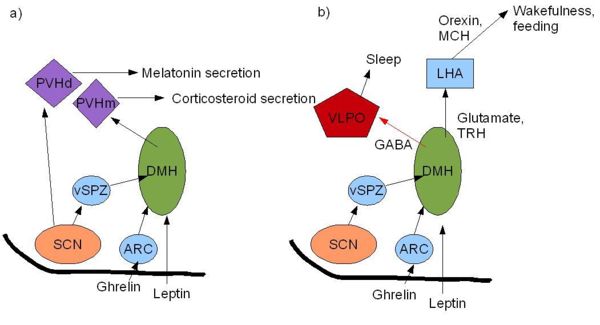
SCN regulation of the sleep/wake cycle; The SCN innervates regions of the brain that regulate the sleep-wake cycle. a) The ventral sub-paraventricular zone (SPZ) projects to the
dorsomedial hypothalamus (DMH) which controls many circadian responses including
corticosteroid secretion. The DMH innervates the medial parvicellular
paraventricular nucleus (PVHm) and regulates the neurons which control corticosteroid production. The SCN also projects to the dorsal parvicellular paraventricular nucleus, (PVHd) which projects to sympathetic preganglionic neurons which regulate
melatonin output from the
pineal gland. b) The DMH also has projections to the ventrolateral preoptic nucleus VLPO (the sleep promoting region) and to the
orexin neurons and
melanin-concentrating hormone (MCH) neurons in the
lateral hypothalamus (LHA) which together regulate sleep and wakefulness. Hormones involved in appetite regulation such as ghrelin and leptin can influence these areas of the brain thus may have a role in resetting the circadian rhythms generated by the SCN. (Adapted from Saper
et al. (2005)
[5])
The circadian clock in mammals sets specific temporal patterns within our bodily systems, including many physiological functions (body temperature, melatonin release, glucocorticoid secretion) and behavioural functions. The "master" circadian clock of mammals is in the suprachiasmatic nucleus (SCN) of the hypothalamus. This receives signals encoding light, which are carried from the retina to the SCN by the retinohypothalamic tract. Vasoactive intestinal polypeptide, vasopressin, somatostatin and other peptides, released from subpopulations of neurones in the SCN, activate and synchronise other SCN neurons, the output of which coordinates behavioural rhythms, including by directing the sleep/wake cycle. The sleep/wake cycle is driven by fluctuations in two main hormones: corticosterone (cortisol in humans) and melatonin. In rats, corticosterone secretion increases during the night when they are active. This rise is followed by an increase in locomotor activity, in which rats forage for food and subsequently begin to feed. Melatonin is an important hormone that is released from the pineal gland during the night and, amongst its many actions, it affects sleep and appetite. However, these hormones have different roles in different species. For example, melatonin is released during the night in both humans and rodents, and yet latter are nocturnal and therefore most active at night. This indicates that melatonin has opposite effects in rodents and humans in that it appears to drive the awake period in rats, but induces sleep in humans. Thus whilst the SCN may regulate the sleep-wake cycle, this regulation is species-specific. Nevertheless, it is possible that the SCN may drive feeding patterns directly as the SCN has reciprocal interactions with orexigenic regions of the brain such as the lateral hypothalamus.
Alternatively, the SCN may initiate feeding by conducting circadian rhythmic oscillations in the hormones involved in appetite. Indeed, a number of hormones involved in feeding behaviour and appetite, including leptin and ghrelin, show circadian oscillations. [6]. Leptin exhibits circadian patterns in both gene expression and protein secretion in humans, with a peak during sleep [7]. In rodents, ablation of the SCN eliminates leptin circadian rhythmicity [8] and yet the role of the SCN in conducting this pattern is unclear.
Whilst the role the SCN has in driving feeding behaviour remains elusive, oscillators in other areas of the brain and other organs may also produce rhythmic patterns in feeding. Rodents show food anticipatory behaviour indicating that these peripheral oscillators can be reset by feeding time itself, thus have been named 'food entrainable oscillators'. It has been suggested that the SCN signals to these other oscillators using signalling molecules such as transforming growth factor alpha (TGF-alpha) and prokinecticin 2 that prevent dampening of circadian rhythms in the tissues [3]. [9]
Peripheral Clocks and Food Entrainable Oscillators
© Image SCN and Peripheral Oscillator Interaction.
The SCN and peripheral clocks are not affected by meal timings, but restriction of food is the dominant synchroniser for peripheral clocks. When rats are restricted of food, metabolic and hormonal factors used by the SCN to drive peripheral oscillators are uncoupled, to shift to food time. Normally, the SCN and peripheral oscillators work together, but a change in food availability can uncouple them when environmental circumstances demand that feeding patterns are shifted from their normal place in the light-dark cycle.[10].
The 'Food Entrained Oscillator' is a mysterious circadian clock, which is independent of the SCN. It ensures the body is ready to digest and extract nutrients from the food in anticipation of meal-time [4]. Clock genes may contribute to the Food Entrained Oscillator, but are not essential – animals without essential clock genes Bmal1 or per1 or per2 were arrhythmic in constant darkness but still could express food anticipation. [11]
Sleep deprivation, shift-work and appetite
In modern society, shopping, eating, working and drinking are widely available 24 hours a day. Over the last few decades, the number of hours of sleep young adults get has decreased by 1-2 hours. This is correlated with the prevalence of obesity within the U.K which has trebled over the last three decades. Although the rise in this obesity epidemic is multi-factorial, sleep deprivation may be another factor to add to the list needing to be addressed in this ever rising health problem. [12] [13] [14].
Sleep deprivation elevates the circulating levels of the appetite stimulating hormone ghrelin and decreases those of leptin[15][14]. These changes could be the causes of increased food intake in those sleep-deprived adults where a rise in body weight is also observed [15]. It is therefore important to understand the health implications that sleep-deprivation, associated with jetlag and shift working, has on the body in order to develop therapeutic schemes against such related disorders as obesity.
Conclusion
Over the last decade, significant progress has been made in our understanding of the inter-relationship between circadian biology, sleep and metabolism in the context of obesity. The SCN, which is a vital coordinator of circadian rhythms, may have a role in driving feeding patterns, along with other peripheral oscillators in the brain and organs that produce rhythmic patterns in feeding time. In the modern world, sleep deprivation is a growing issue of concern and the health implications need to be acknowledged. An adequate number of hours sleep each night is vital for a healthy life. Disruptions of the circadian variations in such appetite-regulating hormones as leptin and ghrelin may contribute to the development of obesity in sleep-deprived patients. Gaps still remain in our knowledge of the underlying biological rhythms and its association with obesity. So far, therapeutic strategies have failed to curb the rising incidence of obesity, but if, through a greater understanding of these complex systems we can control appetite, the benefits will be a healthier population and less strain on the health services.
References
- ↑ Yi CX (2006)Ventromedial arcuate nucleus communicates peripheral metabolic information to the suprachiasmatic nucleus. Endocrinology 147:283–94
- ↑ Schwartz MW (2007)Central nervous system control of food intake Nature 404:661-671
- ↑ 3.0 3.1 3.2 3.3 Froy et al.(2010) Metabolism and circadian rhythms—implications for obesity Endocr Rev 31:1-24
- ↑ 4.0 4.1 Mendoza et al.(2009) Brain clocks: From the suprachiasmatic nuclei to a cerebral network. The Neuroscientist 15:5
- ↑ Saper A et al. (2005) The hypothalamic integrator for circadian rhythms Trends Neurosci 28:152-7
- ↑ Yildiz BO (2004) Alteration in the dynamics of circulating ghrelin, adiponectin and leptin in human obesity Proc Natl Acad Sci USA 101:10434-9
- ↑ Kalra SP(2003) Rhythmic, reciprocal ghrelin and leptin signaling: new insight in the development of obesity Regul Pept 111:1–11
- ↑ Kalsbeek A (2001) The suprachiasmatic nucleus generates the diurnal changes in plasma leptin levels Endocrinology 142:2677–85
- ↑ Gilbert J (2009) Behavioral effects of systemic transforming growth factor-alpha in Syrian hamsters Behav Brain Res 198:440-8
- ↑ Escobar C et al. (2009) Peripheral oscillators are important for food anticipatory activity (FAA) Eur J Neurosci 30:1665–75
- ↑ Pendergast et al' (2009)
- ↑ Gimble JM et al. (2009) Circadian biology and sleep: missing links in obesity and metabolism? Obesity Rev 10(suppl 2):1-5
- ↑ Rennie KL, Jebb SA (2005) Prevalence of obesity in Great Britain Obesity Rev 6:11-2
- ↑ 14.0 14.1 Spiegel et al.(2004) Sleep curtailment in healthy young men is associated with decreased leptin levels, elevated ghrelin levels, and increased hunger and appetite Ann Internal Med 141:846-50
- ↑ 15.0 15.1 Taheri et al. (2004) Short sleep duration is associated with reduced leptin, elevated ghrelin, and increased body mass index PLoS Medicine 1:e62


